
پتانسیل کنش یا پتانسیل عمل (به انگلیسی: Action Potential) فعل و انفعالاتی گذرا در پتانسیل غشاء سلولهای عصبی و عضلانی جهت پیامرسانی در طول شبکه عصبی است.
مراحل اساسی

از لحاظ زمانی، میتوان چهار دوره اساسی را برای یک پتانسیل عمل تعیین کرد:
- تحریک: چنانچه پتانسیل ایجادی از حد آستانه فراتر باشد، سلول تحریک میشود.
- در طی واقطبش، پتانسیل داخل غشاء به دلیل نفوذ یون هایسدیم به سرعت بالا میرود و به قله خود میرسد.
- در طی بازقطبش، پتانسیل عمل کاهش مییابد.
- در دوره تحریک ناپذیری، پتانسیل عمل به مقداراولیه اش بازمیگردد.
در سلولهای عصبی، پتانسیل عمل بر روی غشای تحریک پذیر پدید میآید، طول آکسون را طی میکند و وظیفهٔ پیامرسانی را بر عهده دارد.
جزئیات مراحل پتانسیل
۱- کاهش فعالیت پمپ سدیم/ پتاسیم= کاهش انتقال فعال و هیدرولیز ATP= ابتدای نمودار پتانسیل عمل (هنوز حدود ۶۵-)
۲- باز شدن کانالهای دریچهدار سدیمی 》 ورود سدیم به سلول در جهت شیب غلظت 》 انتشار تسهیل شده 》 مثبت شدن بار سطح داخل غشاء نسبت به خارج آن > بخش بالا رو نمودار پتانسیل عمل (از ۶۵- به سوی ۴۰+) افزایش ۱۰۵ میلی ولت پتانسیل نورون.
۳- باز شدن کانالهای دریچهدار پتاسیمی = خروج پتاسیم از سلول در جهت شیب غلظت = انتشار تسهیل شده = منفی شدن دوباره سطح داخل غشاء نسبت به خارج آن = بخش پایین رو نمودار پتانسیل عمل (از ۴۰+ به سوی کمی کمتر از ۶۵-) (حدوداً ۷۰-) کاهش حدود ۱۱۰ میلی ولت پتانسل نورون.
تذکر: خروج یون پتاسیم نسبت به ورود یون سدیم، بیشتر اختلاف پتانسل را تغییر میدهد.
تذکر: در این حالت با این که بار الکتریکی سطح داخل غشاء نسبت به خارج آن منفی است (کمتر از پتانسیل آرامش است) و غلظت یونها تغییر کرده (این تصور غلط است که فکر کنیم در داخل اکثریت، سدیم و خارج پتاسیم قرار گرفتهاست؛ در هر پتانسیل عمل واحد، تغییر این مقدار سدیم و پتاسیم به قدری کم است که قابل اندازهگیری نیست. در واقع قبل از آنکه غلظت سدیم و پتاسیم آنقدر تغییر کنند که دیگر پتانسیل عملی ایجاد نشود، یک سلول عصبی میتواند ۱۰۰ هزار تا ۵۰ میلیون بار ایمپالس منتقل کند)
۴- فعال شدن دوباره و البته کمی بیشتر از قبل پمپ سدیم-پتاسیم (هیدرولیز ATP) - خروج سدیم از سلول و بازگشت پتاسیم به داخل که باعث افزایش پتانسل -۶۵ در نتیجه حالت آرامش.
پتانسیل عمل در سلولهای عصبی
اطلاعات در طول سلول عصبی به شکل تکانهٔ شیمیائی - برقی حرکت میکند و از مناطق دندریتی به سوی انتهای آکسون میرود. توان ایجاد این پتانسیل عمل مختص سلولهای عصبی و ناشی از انبوه مجراهای یونی (ion channels) و تلمبههای یونی (ion pumps) موجود در غشاء یاخته است. مجراهای یونی همان مولکولهای پروتئینی حلقوی است که منفذهای غشاء سلول عصبی را میسازند. این ساختارهای پروتئینی از راه باز و بسته کردن منفذها به تنظیم مبادلهٔ یونهای دارای بار الکتریکی نظیر سدیم (+Na) و پتاسیم (+K) و کلسیم (++Ca) یا کلر (-Cl) میپردازند، در تمام مدت پتانسیل آرامش یا پتانسیل عمل، هم خروج و هم ورود مواد به داخل سلول از طریق کانالهای نشتی امکانپذیر میشود و فقط وابسته به کانالهای دریچهدار نمیباشد؛ هر مجرای یونی (چه نشتی و چه دریچهدار) به صورت انتخابی عمل میکند و معمولاً به یون معینی اجازهٔ عبور میدهد. ساختارهای پروتئینی (تلمبههای یونی) جداگانهای از راه تلمبه کردن یونهای گوناگون به درون یا بیرون نورون، آنها را در دو سوی غشاء یاخته با توزیعی نامتوازن نگه میدارند. تلمبهٔ یونی از همین راه در شرایط استراحت نورون، تراکم زیاد +Na را در بیرون و تراکم اندک آن را در درون نورون حفظ میکند. اثر کلی تلمبهها و مجراهای یونی، قطبیسازی غشاء نورون است به طوری که بار الکتریکی (charge) مثبت در بیرون غشاء و بار الکتریکی منفی در درون آن میماند.
هرگاه نورون در حال استراحت تحریک شود، اختلاف ولتاژ دو سوی غشاء آن کاهش مییابد. اگر پتانسیل به اندازهٔ کافی کم شود، مجراهای +Na در نقطهٔ تحریک برای مدت کوتاهی باز شده و یونهای سدیم به درون یاخته سرازیر میشوند. نام این فرایند ناقطبیشدن (depolarization) است. در این شرایط، بار الکتریکی درون آن ناحیه از غشاء یاخته نسبت به بیرون آن مثبت میشود. مجراهای +Na بعدی کاهش ولتاژ را درمییابند و باز میشوند و در نتیجه منطقهٔ مجاور ناقطبی میشود. مجراهای +Na بعدی آن نیز کاهش ولتاژ را درمییابند و باز میشوند و در نتیجه منطقهٔ مجاور بعدی ناقطبی میشود. این فرایند خودنگهدارندهٔ (self - propagating) ناقطبیشدن (که در طول جسم یاخته تکرار میشود) تکانهٔ عصبی را شکل میدهد. همینکه تکانهٔ عصبی از نقطهای میگذرد مجراهای +Na بسته شده و تلمبههای یونی گوناگونی فعال میشوند تا غشاء یاخته را به سرعت به حالت استراحت برگردانند.
حرکت پتانسیل عمل در نورونها
سرعت تکانهٔ عصبی هنگام سفر به انتهای آکسون، بسته به قطر آکسون، بین ۳ تا ۳۰۰ کیلومتر در ساعت است. این سرعت عموماً در آکسونهای بزرگتر بیشتر است. این سرعت به میلینپوش بودن یا نبودن آکسون نیز بستگی دارد. در غلاف میلین، یاختهای تخصصیافتهٔ گلیائی یکی پس از دیگری خود را به آکسون پیچیدهاند و بین هر دو یاختهٔ گلیائی فرورفتگی کوچکی هست. خاصیت عایقسازی غلاف میلین موجب میشود تکانهٔ عصبی در واقع از بندی به بند دیگر بپرد و از این راه سرعت انتقال را بسیار زیاد کند. غلاف میلین (جنس آن از فسفولیپید و پروتئین است که توسط سلولهای پشتیبان ساخته میشود، توجه شود که بسیاری از سلولهای عصبی دارای میلین هستند نه همهٔ آنها) از اختصاصات جانوران ردههای بالای تکاملی است، و بهویژه در مناطقی از دستگاه عصبی بیشتر است که سرعت انتقال در آنها بسیار مهم باشد. بیماری اسکلروز چندگانه (multiple sclerosis) (تصلب چندگانه) که اختلال شدید اعصاب حسی و حرکتی از ویژگیهای آن است، ناشی از تباهی همین غلاف میلین است.
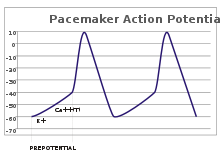
پتانسیل عمل در سلولهای قلبی
در سلولهای قلبی بهخصوص در گرههای قلبی که تولیدکننده ضربان هستند نیز پتانسیل عمل تولید میشوند. در سلولهای عضلانی قلب پتانسیل عمل تولید شده دارای فاز کفه است.
جستارهای وابسته
- نورون
- در بحث کدگذاری محتوای اطلاعات در مغز، پتانسیل عمل بیشتر با عنوان شلیک (علوم اعصاب) عنوان میشود.
- آرتور گایتون، جان ادوارد هال (۱۳۸۶)، «فیزیولوژی عصبی»، فیزیولوژی پزشکی گایتون، ترجمهٔ احمدرضا نیاورانی، سماط، شابک ۹۶۴-۵۷۸۹-۹۸-۲
- تونی اسمیت، سو دیویدسن (۱۳۸۴)، مغز و سیستم عصبی، ترجمهٔ عباس تیرگانی، بیژن معصوم، نشر سنبله، شابک ۹۶۴-۳۹۲-۰۷۱-۲