
بازسازی دیانای به مجموعهای از فرایندهای مولکولی گفته میشود که در آن یک سلول شناسایی و آسیب وارد شده به دیانای آن اصلاح میگردد. به عبارت دیگر، در فرایند بازسازی دیانای، ژنوم سلول مجدداً کدگذاری میشود. در سلولهای انسان، هم فعالیتهای متابولیکی طبیعی و هم فاکتورهای زیستمحیطی مانند نور و تشعشع فرابنفش میتواند باعث آسیب به دیانای شود. آسیب ژنوم سلولهای انسان به میزان ۱ میلیون ضایعه مولکولی به ازای هر سلول در هر روز رخ میدهد. بیشتر این آسیبها مربوط به ساختار مولکول دیانای است و میتواند توانایی سلول را برای رونویسی ژن مؤثر در کدگذاری دیانای تحت تأثیر قرار دهد. آسیبهای دیگر به صورت بالقوه منجر به ایجاد جهشهای مضر در ژنوم سلولی میشوند که بر روی زنده ماندن سلولهای دختر بعد از رشتمان اثر میگذارد. فرایند بازسازی دیانای به صورت دائمی فعال است و به آسیب وارد شده بر ساختار دیانای پاسخ میدهد. زمانی که فرایند بازسازی نرمال دچار مشکل شده و خزان یاختهای رخ نمیدهد، این امکان وجود دارد که دیانای آسیب دیده و بازسازی ناپذیری ایجاد شود. این وضعیت با شکستگیها در رشته دوگانه و هم بری یا اتصال عرضی دیانای همراه خواهد بود. پیوند اشتراکی بین دو تیمین مجاور نتیجه این جهش است. (اتصال عرضی بین رشتهای یا ICL
میزان بازسازی دیانای بستگی به فاکتورهای زیادی دارد که شامل نوع، سن و محیط درون سلول میباشد. یک سلول ممکن است شامل مقدار زیادی از دیانای های آسیب دیده یا به صورت بازسازیهای نامؤثری باشد. چنین شرایطی میتواند در یکی از سه حالت زیر رخ دهد:
- حالت برگشتناپذیر خفتگی یا کمون که به صورت فرایند پیری شناخته شدهاست.
- زیان رساندن سلول به خود (خودکشی سلول) که تحت عنوان خزان یاختهای یا مرگ برنامهریزی شده سلولی شناخته میشود.
- تقسیم سلولی نامنظم که میتواند منجر به شکلگیری توموری شود که سرطان نام دارد.
توانایی بازسازی دیانای در یک سلول برای یکپارچگی ژنوم و بنابراین برای عملکرد طبیعی ارگانیسم نیز حیاتی است. بیشتر ژنها به صورت ابتدایی مؤثر بر طول عمر نشان داده شدهاست که بر روی بازسازی دیانای آسیبدیده و حفاظت از آن تأثیر دارد.
جایزهٔ نوبل ۲۰۱۵ شیمی به منظور کار بر روی سازوکار مولکولی فرایندهای بازسازی دیانای، به توماس لیندال، پال مودریچ و عزیز سنجر اعطا گردید.
آسیب به دیانای
دیانای آسیبدیده ناشی از فاکتورهای زیستمحیطی و فرایندهای متابولیک طبیعی در درون سلول، به میزان ۱۰۰۰۰ تا ۱۰۰۰۰۰۰ آسیب و زخم مولکولی به ازای هر سلول در هر روز اتفاق میافتد. در حالی که این تنها شامل ۰۰۰۱۶۵/۰ درصد از تقریباً ۶ بیلیون ژنوم بازی (۳ بیلیون جفت باز) است، زخمهای (آسیبهای) بازسازی نشده در ژنهای حیاتی (مانند ژن سرکوبگر غده) میتوانند مانع توانایی سلول برای اجرای عملکردشان و افزایش ارزیابی احتمال شکلگیری تومور و مشارکت در ناهمسانی توموری شود.
اکثر دیانایهای آسیب دیده بر روی ساختار اولیه هلیکس دوگانه اثر میگذارند که در این صورت بازها خودشان از نظر شیمیایی تغییر مییابند. این تغییرات میتواند در ابتدا منجر به تخریب ساختار مارپیچی منظم مولکولی با معرفی پیوندهای شیمیایی غیر وابسته به مکان خاصی یا ترکیبهای افزایشی حجیم شود که متناسب با ساختار مارپیچی دوگانه استاندارد نیست. برخلاف پروتئینها و آرانای، دیانای معمولاً فاقد ساختار جابهجاسازی سه اتم یا بنیان است و بنابراین آسیب یا فروپاشی در این سطح رخ نمیدهد. به هر حال دیانای فوق مارپیچ و زخم در اطراف پروتئینهای بستهبندیکننده، هیستون نام دارد (در یوکاریوتها) و هر دو ساختار فوق به اثرات مربوط به آسیب دیانای آسیبپذیر هستند.
آسیب دیانای میتواند به دو نوع اصلی تقسیم گردد:
- آسیبهای درونزاد مانند حمله گونههای دارای اکسیژن فعالشده که توسط محصولات جانبی متابولیک نرمال تولید میشود (جهش خود به خودی)، به ویژه میتوان به فرایند ازدستدادن آمیناکسیداتیو اشاره کرد.
- همچنین شامل تکرار خطا نیز میشود.
- آسیب برون زا که توسط عوامل خارجی به وجود میآید و عبارتند از:
تکرار دیانای آسیب دیده قبل از تقسیم سلولی میتواند منجر به درآمیختن غلط بازهای مخالف که آسیب زا هستند، شود. سلولهای دختری که از این بازهای غلط به ارث رسیدهاند جهشهایی را انتقال میدهند که از توالی اصلی دیانای غیرقابل جبران هستند (به جز در مورد نادری که به عنوان مثل جهش رو به عقب از طریق تبدیل ژن صورت میگیرد).
انواع آسیبها
چندین نوع آسیب به دیانای وجود دارد که ناشی از فرایندهای سلولی داخلی هستند:
- آلکیلدار کردن بازها (برای مثال ۸- اکسو- ۷ و ۸- دی هیدروگانین (8-oxoG)) و وقفه در تولید رشته دیانای از گونههای اکسیژن فعال شده
- متیلدار کردن بازها (معمولاً متیلی شدن) مانند شکلگیری ۷- متیل گانین، ۱- متیل آدنین، ۶-او-متیلگوانین
- آبکافت بازها مانند از دست دادن گروه آمین، دیآمیناسیون و دیپوریاسیون.
- تشکیل ترکیبات ورینی بزرگ (به عنوان مثال بنزو (آ) پیرین دی ال اپوکسید- dG ورینی، آریستولاکتام I-dA ورینی)
- جور شدن اشتباه بازها به دلیل خطاها در همتاسازی و همانندسازی دیانای که در این صورت باز اشتباه دیانای در مکان شکلگیری رشته جدید دیانای قرار میگیرد یا این باز دیانای از مکان مورد نظر خارج میشود.
- آسیب تک ورینی که منجر به ایجاد تغییر در تک بازهای نیتروژنی دیانای میشوند.
- آسیب دی ورینی
آسیب ایجاد شده توسط عوامل خارجی به شکلهای مختلفی بروز میکند. برخی از مثالها در این زمینه عبارتند از:
- نور UV-B که منجر به شکلگیری پیوندهای غلط بین سیتوزینهای مجاور و بازهای تیامین میشود که ایجادکننده دیمرهای پیریمیدین است. این حالت آسیب مستقیم به دیانای نام دارد.
- نور UV-A به مقدار زیادی رادیکالهای آزاد تولید میکند. آسیب ایجاد شده توسط رادیکالهای آزاد آسیب غیر مستقیم به دیانای نام دارد.
- پرتو یونی مانند آنچه توسط تجزیه رادیو اکتیو یا در پرتوهای کیهانی رخ میدهد که میتواند باعث شکستن رشتههای دیانای شود. اشعه یونیزاسیون در سطح کم ممکن است ایجادکننده آسیبهای جبرانناپذیری به دیانای باشد (منجر به ایجاد خطاهای همتاسازی و نسخهنویسی میشود که برای نئوپلازیا مورد نیاز است یا برای تداخلات ویروسی نیاز است) که منجر به پیری قبل از رسیدن به بلوغ یا سرطان میشود.
- اختلال حرارتی در مقادیر بالای دمایی منجر به افزایش دیپیوریاسیون (از دست دادن بازهای پورین از چارچوب اصلی دیانای) و شکستن رشتههای مجزا و تکی میشود. به عنوان مثال هیدرولیز دی پیوریلیشن در باکتری گرمادوست مشاهده شدهاست که در چشمههای آب گرم در دمای ۴۰ تا ۸۰ درجه سلسیوس رشد میکنند. میزان دی پیوریشن یا تخریب بازهای پورین (۳۰۰ پورین باقیمانده به ازای هر ژنوم به ازای هر نسل) در این گونهها بالا است که توسط مکانیسم بازسازی نرمال بازسازی میگردد و بنابراین احتمالاً امکان رد کردن رفتار ناسازگارانه وجود ندارد.
- مواد شیمیایی صنعتی مانند وینیل کلرید و هیدروژن پراکسید و مواد شیمیایی زیستمحیطی مانند هیدروکربنهای آروماتیک چندحلقهای یافت شده در دود و دوده میتوانند ورینهای دیانای گوناگون، اتانوبازها، بازهای اکسید شده، فسفوتری استرهای قلیایی شده و دیانای و دیانای کراس لینکینگ را ایجاد نمایند.
آسیب UV، قلیایی شدن، متیلی شدن، آسیب اشعه ایکس و آسیب اکسیدی شدن مثالهایی از آسیبهای ذکر شده هستند. آسیب خود به خودی میتواند شامل از دست دادن باز، دامانسیون، شکستن حلقه شکر و تغییر در تاتومریک باشد.
آسیب هستهای در مقابل آسیب به دیانای میتوکندری
در سلولهای انسان، و در سلولهای یوکاریوتی به صورت کلی، دیانای در دو موقعیت سلولی در درون هستهها و در درون میتوکندری یافت میگردد. دیانای هسته یا nدیانای به صورت کروماتین در مراحل غیر همانند چرخه سلولی وجود دارد و به صورت ساختار دانهای متراکم تحت عنوان کروموزومها در طول تقسیم یاخته متراکم شدهاست. در هر مرحله، دیانای به مقدار زیادی فشرده میگردد و در اطراف پروتئینهای مهره مانند به نام هیستونها چرخش مییابد. در هر جایی که سلول نیاز به بیان اطلاعات ژنتیکی کدگذاری شده در nدیانای خودش داشته باشد در این صورت ناحیه کروموزومی مورد نیاز از هم میپاشد، ژنهای واقع بر روی آن بیان میگردند و سپس ناحیه به ساختار استراحت خود به حالت متراکم بازمیگردد. دیانای میتوکندری یا mtدیانای درون اندامکهای میتوکندری قرار دارد، که از آن چندین کپی وجود دارد و علاوه بر این به مقدار زیادی در ارتباط با تعدادی از شکلهای پروتئینی پیچیده است که تحت عنوان نوکلوئید شناخته شدهاند. درون میتوکندری، گونههای اکسیژن رآکتیو یا ROS یا رادیکالهای آزاد محصولات جانبی تولید ثابت آدنوزین تریفسفات یا ATP از طریق فسفرگیری اکسایشی هستند که ایجاد محیطی اکسیداتیو میکنند که برای آسیب به mtدیانای شناخته شدهاست. آنزیم اصلی برای مقابله با سمیت این گونهها سوپر اکسید دیسموتاز است که هم در میتوکندری و هم در سیتوپلاسم سلولهای یوکاریوتی وجود دارد.
پیری و آپوپتوز
پیری یک وضعیت غیرقابل برگشت است که تقسیم سلول و پاسخ حفاظتی به کوتاه شدن کروموزوم به پایان میرسد. تلومرها نواحی طولانی از دیانای غیرکدگذاریشده تکراری هستند که کروموزومها را پوشش میدهند و در نهایت تخریب جزئی مربوط به تقسیم سلولی در هر زمان را تحت کنترل دارند (حد هایفلیک را ببینید). برعکس، خاموشی یک حالت برگشتپذیر برای خواب سلولی است که در ارتباط با آسیب ژنومی نیست (چرخه یاختهای را ببینید). پیری در سلولها ممکن است به عنوان تغییر عملکردی برای آپوپتوز در مواردی عمل نماید که حضور فیزیکی سلول برای دلایل مکانی و فضایی به وسیله ارگانیسم مورد نیاز است که به صورت مکانیسم بازسازی نهایی برای جلوگیری از آسیب تکرار شدن نامناسب دیانای در غیاب رشد نشانهگذاری یاخته به کار برده میشود. تقسیم سلولی بی نظم میتواند منجر به شکلگیری تومور شود (سرطان را ببینید) که به صورت بالقوهای برای ارگانیسم کشنده است؛ بنابراین القا پیری و آپوپتوز به عنوان بخشی از استراتژی حفاظت در مقابل سرطان در نظر گرفته میشود.
آسیب دیانای و جهش
تشخیص میان دیانای آسیب دیده و جهش مهم است، این دو حالت دو نوع اصلی از خطا در دیانای هستند. آسیبهای دیانای و جهش به صورت اساسی با هم متفاوت هستند. آسیبها به صورت وضعیتهای فیزیکی غیرطبیعی در دیانای هستند مانند شکستن رشته دوتایی یا مجزا، بقایای ۸-اوکسو-۲-دئوکسی گوآنوزین و مواد افزودنی هیدرو کربنهای آروماتیک پلی سایکلیک. آسیبهای دیانای میتواند به وسیله آنزیمها شناسایی گردد و بنابراین آنها میتوانند به درستی بازسازی گردند در صورتی که اطلاعات اضافی مانند توالی غیر آسیب دیده و سالم در رشته مکمل دیانای یا در کروموزوم همولوگ برای کپی کردن در دسترس باشند. اگر سلول آسیب دیده دیانای حفظ گردد، در این صورت مانع از نسخهنویسی ژن میشود و بنابراین ترجمه به پروتئین نیز مسدود میگردد. تکرار نیز ممکن است مسدود گردد یا سلول ممکن است بمیرد.
برعکس دیانای آسیب دیده، جهش تغییر در توالی بازی دیانای است. جهش نمیتواند با کمک آنزیمها در زمان وجود تغییر بازی در هر دو رشته دیانای تشخیص داده شود و بنابراین جهش قابل بازسازی یافتن نیست. در سطح سلولی، جهشها میتوانند باعث تغیر در عملکرد پروتئین و نظم آنها شوند. جهشها در زمانی تکرار میشوند که سلول تکثیر میگردد. در سلولها، سلولهای جهش یافته به صورت مکرر در حال افزایش یا کاهش بر طبق اثرات جهش روی توانایی سلول به منظور زنده ماندن یا تکثیر هستند. اگر چه به صورت مطلق با یکدیگر تفاوت دارند، دیانای آسیب دیده یا جهش یافته به هم مرتبط هستند زیرا دیانایهای آسیب دیده اغلب منجر به ایجاد خطاهایی در سنتز دیانای در طول فرایند تکثیر یا بازسازی میشوند، این خطاها منبع اصلی جهش هستند.
این مشخصات هم برای دیانای آسیب دیده و هم جهش یافته مشخص شدهاند که میتوان در آسیبهای دیانای مشاهده کرد که مسئلهای خاص در زمینه عدم تقسیم شدن یا تقسیم شدن آهسته سلولها به حساب میآید که در این صورت آسیبهای بازسازی نیافته میتواند در طول زمان تجمع یابند. از سوی دیگر در تقسیم سلولی سریع، دیانایهای آسیب دیده و بازسازی نیافته به نابودی سلول با استفاده از مسدود ساختن تکثیر نمیپردازند بلکه منجر به ایجاد خطاهایی در تکثیر و بنابراین جهش میشوند. اکثر جهشها دارای تأثیری خنثی و بیاثر بر روی بقای سلول و زنده ماندن آنها نیستند؛ بنابراین در سلولهای یک بافت دارای تکثیر سلول، سلولهای جهش یافته تمایل به از بین رفتن دارند. به هر حال جهشهای نادری ممکن است منجر به ایجاد برتری زنده ماندن با توجه به کلونی توسعه یافته در مجاورت سلولهای بافت شود. این مزیت برای یک سلول به عنوان عیبی برای کل ارگانیسم بهشمار میآید زیرا چنین سلولهای جهش یافتهای میتواند منجر به ایجاد سرطان شوند؛ بنابراین دیانایهای آسیب دیده در اثر تقسیم سلولیهای مکرر منجر به افزایش جهشها میشوند و در نتیجه سرطان شکل میگیرد. برعکس، دیانایهای آسیب دیده در تقسیم سلولی نادر میتواند منجر به ایجاد پیری زودهنگام شود.
مکانیسمهای بازسازی دیانای
سلولها در صورتی که دیانای آسیب دیده منجر به از دست دادن یکپارچگی و عدم دسترسی به اطلاعات ضروری ژنوم گردد نمیتوانند فعالیت نمایند (اما بقیه سلول در زمانی که ژنهای غیر اساسی از بین میروند یا آسیب میبینند تا حدی به فعالیت میپردازند). بسته به نوع آسیب وارد شده به ساختار دوگانه مارپیچ دیانای، روشهای بازسازی مختلفی برای حفظ بقیه اطلاعات به کار برده میشود. در صورت امکان، این سلولها از رشته مکمل تغییر نیافته دیانای یا کروماتید خواهری به عنوان الگویی برای بازیابی اطلاعات اولیه استفاده میکنند. بدون دسترسی به الگو، سلولها از مکانیسم بازیابی خطای به عنوان سنتز ترنسلیژن یا ترجمه آسیب و زخم به عنوان آخرین چاره استفاده میکنند.
آسیب به دیانای شکلگیری فضایی مارپیچ را تغییر میدهد و چنین تغییراتی میتواند به وسیله سلول نشان داده شود. در ابتدا آسیب دیانای به صورت موضعی تعیین میگردد، مولکول های خاص برای بازسازی دیانای به آن یا در نزدیکی مکان آسیب اتصال مییابند که منجر به القای اتصال مولکولهای دیگر و ایجاد ساختار پیچیدهای میشود که منجر به ایجاد بازسازی حقیقی و واقعی میگردد.
برگشت مستقیم
سلولهایی که محدودکننده این سه نوع آسیب به دیانای هایشان توسط برگشت شیمیایی آن هستند شناخته شدهاند. این مکانیسمها نیازی به بازسازی نمونه ندارد زیرا انواع آسیبهایی که آنها میتوانند ایجاد نمایند تنها در یکی از چهار باز رخ میدهد. چنین مکانیسمهای برگشت مستقیم در ارتباط با نوع آسیب وارده و شکستگی پایه فسفو دی استر ربطی ندارد. شکلگیری دایمرهای پیریمیدین بر اثر اشعه UV منجر به ایجاد پیوندهای کووالانسی غیرطبیعی بین بازهای پیریمیدن جفت شده میشود. فرایند فوتونی اکتیویشن (فعال شدن در اثر نور) به صورت مستقیم منجر به برگشت آسیب توسط فعالیت آنزیم فوتولیاز میشود که کار فعال ساختن را به صورت اجباری و ناخواسته انجام میدهد و این فعالیت بستگی به انرژی جذب شده توسط نور آبی UV برای انجام کاتالیز دارد (طول موج ۳۰۰ تا ۵۰۰ نانومتر). نوع دیگر آسیب مربوط به متیلاسیون باز گوانین است که به صورت مستقیم توسط پروتئین متیل گوانین متیل ترنسفراز یا MGMT معکوس میگردد و معادل باکتریایی آن ogt نامیده میشود. این یک فرایند پر هزینهای است زیرا هر مولکول MGMT میتواند تنها برای یک بار مورد استفاده قرار گیرد از این رو واکنش در مقایسه با فرکافتی به صورت استوکیومتری است. پاسخ کلی به عوامل متیلکننده در باکتری به صورت پاسخ سازگارانه نشان داده شدهاست و اشاره به سطح مقاومت عوامل قلیایی یا آلکیلهکننده در معرض تنظیم ارتقا یافته آنزیمهای بازسازی آلکیل دار شدن مینماید. نوع سوم از دیانای آسیب دیده در ارتباط با سلولهایی است که دارای متیلاسیون معینی برای بازهای سیتوزین و آدنین هستند.
آسیب تک رشتهای

زمانی که تنها یکی از دو رشته مارپیچی دوگانه دچار مشکل میشوند، رشته دیگر میتواند به عنوان یک الگو به منظور هدایت صحیح رشته مورد نیاز به کار برده شود. به منظور بازسازی آسیب یکی از دو مولکول جفت شده دیانای، تعدادی مکانیسمهای بازسازی برش وجود دارد که نوکلئوتید آسیب دیده را حذف مینماید و آن را با نوکلئوتید سالم جایگزین کرده و به این شکل رشته دیانای سالم را به وجود میآورد.
- بازسازی برش بازی (BER) که به بازسازی تک باز آسیب دیده حاصل از اکسیداسیون، قلیایی شدن، هیدرولیز یا دامیناسیون میپردازد. باز آسیب به وسیله دیانای گلیسولاز حذف میگردد. دندانه گم شده سپس توسط آنزیمی تشخیص داده میشود که ایپی اندونوکلئوآز نام دارد که پیوند فسفودیاستر را میشکند. بخش گم شده سپس مجدداً به وسیله دیانای-پلیمراز و آنزیم دیانای لیگاز سنتز میگردد و بخش نهایی را شکل میدهد.
- بازسازی برش نوکلئوتید (NER) که ضایعات مارپیچ انحرافی مانند دیمرهای پیریمیدین و ۶ و ۴- محصولات نوری را شناسایی مینماید. شکل خاصی از NER به عنوان بازسازی- رونویسی آنزیمهای NER نسبت به ژنهایی شناخته شدهاست که به صورت فعال قابل رونویسی هستند.
- بازسازی اشتباه جفت شده (MMR) که خطاهای مربوط به همانندسازی دیانای و نوترکیبی را تصحیح میکند که منجر به ایجاد جفت شدنهای اشتباه نوکلئوتیدها میشود (اما بدون آسیب است).
شکستگیهای رشته دوتایی
شکستگیهای رشته دوتایی که در آن هر دو رشته در یک مارپیچ دوگانه هستند و به صورت خاصی به سلول آسیب وارد میکنند زیرا منجر به شکلگیری چیدمان مجدد ژنوم میشود. سه مکانیسم موجود برای بازسازی بریکها یا شکستگیهای رشته دوگانه (DSB) وجود دارد که عبارتند از: اتصال پایانی غیر همولوگی (NHEJ)، اتصال پایانی سطح میکرو همولوژی (MMEJ) و ترکیب مجدد همولوگها. PVN Acharya اشاره به شکستگیهای رشته دوگانه و اتصال متقاطع هر دو رشته در نقطه یکسانی دارد که جبران پذیر نیست زیرا هر رشته میتواند به عنوان الگویی برای بازسازی مورد استفاده قرار گیرد. سلول در میتوزهای بعدی میمیرد یا این که در برخی موارد نادر جهش مییابد.
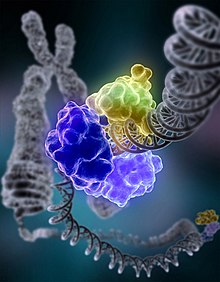
در NHEJ، آنزیم دیانای لیگاز ۴ که به صورت آنزیم دیانای لیگاز خاصی است ترکیبی را با کوفاکتور XRCC4 شکل میدهد که به صورت مستقیم به دو انتها اتصال مییابد. به منظور راهنمایی برای بازسازی دقیق، NHEJ به توالی همولوگی کوتاهی اشاره دارد که میکرو همولوگها نام دارد که در دنبالههای رشتهای تکی در انتهای دیانای برای اتصال وجود دارد. اگر این این مشکلات هم سو هستند در این صورت بازسازی معمولاً دقیق است. NHEJ نیز میتواند جهشهایی را در طول بازسازی معرفی نماید. از دست دادن نوکلئوتیدهای آسیب دیده در مکان شکستن میتواند منجر به حذف و اتصال شکلهای غیر منطبق تغییر مکان یافته شود. NHEJ به صورت خاصی قبل از زمان تکثیر شدن سلول در دیانای خودش دارای اهمیت است زیرا هیچ گونه الگوی در دسترسی برای بازسازی به وسیله ترکیب مجدد همولوگها وجود ندارد. نسخه پشتیبانی برای مسیرهای NHEJ در یوکاریوتهای عالی تر وجود دارد. به غیر از نقش آن به عنوان ژنوم سرپرست، NHEJ نیاز به اتصال هپارین به رشته دوگانه دارد که در طول ترکیب مجدد V(D)J شکل میگیرد، این فرایند ایجادکننده گوناگونی در سلول بی و گیرنده لنفوسیت تی در دستگاه ایمنی مهرهداران است.
ترکیب مجدد همولوگها نیاز به حضور توالی اتفاقی نزدیک به هم و منطبق دارد که به عنوان الگویی برای بازسازی شکستگی مورد استفاده قرار میگیرد. پاسخ خودکار آنزیمی برای این فرایند بازسازی تقریباً مساوی با پاسخ خودکار برای کراسینگاور کروموزومها در میوز است. این مسیر به کروموزومهای آسیب دیده اجازه بازسازی را با کمک کروماتید خواهری (در G2 بعد از تکثیر دیانای در دسترس است) یا کروموزوم همتا به عنوان الگو را میدهد. DSBها منجر به تکثیر خودکار فرایند سنتز در طول رشته شکسته مجزا یا آسیب بازسازی نیافته حاصل از فروپاشی دوشاخه همتاسازی میشوند و اساساً به وسیله ترکیب شدن مجدد بازسازی مییابند.
توپوایزومرازها هم در شکستگیهای تک رشتهای و هم دوتایی در زمان تغییر حالت دیانای سوپرکولینگ معرفی شدهاند که به صورت خاصی در نواحی نزدیک به دوشاخه همتاسازی باز رایج است. چنین شکستگیهایی به صورت دیانای آسیب دیده در نظر گرفته نمیشوند زیرا آنها سطح مشترک طبیعی در توپوایزومرازها با مکانیسم بیوشیمیایی هستند و فوراً به وسیله آنزیمهایی بازسازی مییابند که آنها را ایجاد کردهاند.
تیمی از محققان فرانسوی به مطالعه مکانیسم بازسازی دیانای با رشته دوگان شکسته در موجود زنده پرداختهاند. حداقل دو کپی از ژنوم با شکستگیهای تصادفی دیانای میتواند بخشهای دیانای را از طریق شکلگیری مجدد شکل دهد. به صورت خاصی بخشهای هم پوشانیکننده سپس برای سنتز نواحی همساختی از طریق حرکت حلقه دی مورد استفاده قرار میگیرند که میتواند برای گسترش دادن ادامه یابد تا زمانی که آنها رشتههای مکمل همتایی را بیابند. در مرحله نهایی متقاطع به وسیله RecA وابسته به ترکیب مجدد یا نوترکیبی همولوگی وجود دارد.
سنتز ترانسلسیون
سنتز ترانسلسیون یا TLS به صورت فرایند مقاوم به آسیب دیانای است که به مکانیسم تکثیر دیانای اجازه همانندسازی زخمهای قبلی دیانای را مانند دیمرهای تیامین یا مکانهای ایپی را میدهد. این شامل سوئیچینگ منظم پلیمرازهای دیانای برای ترنسلیشن پلیمرازهای خاص است که اغلب با مکانهای فعال بزرگتری در ارتباط است که میتواند جفت شدن بازهای مخالف مربوط به نوکلئوتیدهای آسیب دیده را فراهم سازد. پلیمراز سوئیچینگ به عنوان واسطهای در بین فاکتورهای دیگر عمل میکند، اصلاح معمولاً بعد از ترجمه و رونویسی فاکتور تکثیر PCNA صورت میگیرد. پلیمراز سنتز ترنسلیژن اغلب دارای اتصال سستی براساس الگوهای سالم مرتبط با پلیمرازهای منظم است. به هر حال بیشتر آنها به مقدار زیادی بر روی بازهای صحیح و متضاد با انواع خاصی از آسیبها تأثیر دارند. به عنوان مثال Pol η به عنوان سطحی عاری از خطا برای جبران زخمهای ایجاد شده توسط اشعه فرابنفش است در حالی که Pol ι جهشهایی را در این مکانها ایجاد میکنند. Pol η به عنوان یک عامل اضافهکننده به آدنین در راستای فوتودیمر T^T با استفاده از جفت شدن بازی واتسون- کریک شناخته شدهاست و آدنین ثانویه به شکلگیری syn آن با استفاده از جفت باز هاگستین اضافه میگردد. از نظر سلولی، خطر مربوط به معرفی جهشهای نقطهای در طول سنتز ترنسلیژن ممکن است برای بازسازی مجدد نسبت به بیشتر مکانیسمهای قوی تر برای بازسازی دیانای ترجیح داده شود که در این صورت ممکن است باعث تودهای شدن کروموزومی یا مرگ سلولی شود. بهطور مختصر، فرایند شامل پلیمراز شدن خاصی است که هر یک به جبران یا بازسازی زخمها در مکانهایی میپردازد که محل تکثیر دیانای است. به عنوان مثال پلیمراز دیانای انسانی اتا میتواند با زخمهای دیانای پیچیدهای مانند جهشهای شبه هدف بازسازی گردد. پارومیتا راچادهاری و آشیس باسو سمیت و جهش زا بودن زخم یکسانی را در اشرشیا کولای با استفاده از تکثیر G[8,5-Me]T تغییر دهنده پلاسمید در اشرشیا کولای با پلیمراز خاص دیانای مورد مطالعه قرار دادند. قابلیت زیستن در رشته فاقد pol II, pol IV و pol V بسیار کم بود، سه SOS محرک پلیمرازهای دیانای نشان دادند که سنتز ترسلیژن با کمک این پلیمرازهای خاص دیانای صورت میگیرد. پلت فرم بای پس برای این پلیمرازها توسط تکثیر آنتی ژن هستهای سلول یا PCNA فراهم میگردد. تحت شرایط نرمال، PCNA پلیمرازهای تکثیرکننده دیانای اتصال مییابد. در مکان زخم، PCNA با کمک پروتئینهای RAD6/ RAD18 به منظور فراهم ساختن پلت فرمی برای پلیمرازهای خاص بای پس زخم و تکثیر دیانای تغییر مییابد. بعد از سنتز ترنسلیژن، اکستانسیون مورد نیاز است. این اکستانسیون یا کشش میتواند با کمک پلیمرازهای تکرار شونده صورت گیرد در صورتی که TLS عاری از خطا باشد، چنانچه در مورد Pol η، اگر TLS حاصل انطباق و اتصال اشتباه باشد در این صورت پلیمراز خاصی برای ادامه دارد آن مورد نیاز است که آن Pol ζ است. Pol ζ منحصر به فرد است و میتواند در انطباقهای اشتباه در بخش پایانی گسترش یابد در حالی که بیشتر پلیمرازهای دیگر توانایی این کار را ندارند؛ بنابراین در زمانی که زخم رخ میدهد، دو شاخه تکثیر مشغول فعالیت خواهد شد، PCNA از پلیمراز processiveبه سمت پلیمراز TLS مانند Pol ι به منظور بازسازی زخم راهاندازی میگردد، سپس PCNA ممکن است به راهاندازی Pol ζ برای گسترش دادن عدم مطابقت میپردازد و PCNA نهایی نسبت به پلیمراز processive راهاندازی خواهد شد.
پاسخ سراسری و کلی به دیانای آسیب دیده
سلولهایی که در معرض تشعشع یونیکننده، نور فرابنفش یا مواد شیمیایی قرار دارند مستعد برای ایجاد ضایعات مختلف دیانای و شکستگیهای دو رشته میباشد. به هر حال، عوامل آسیبرسان به دیانای میتواند دیگر زیستمولکولها مانند پروتئینها، کربوهیدراتها، لیپیدها و RNA باشد. به صورت خاصی با تجمع آسیب، شکستگیهای رشته دوگانه یا ترکیبات افزوده شده به دوشاخههای همتاسازی یا تکثیر سیگنالهای تحریک شناخته شدهای را برای پاسخ کلی به آسیب دیانای تولید میکنند. پاسخ کلی به آسیب به صورت فعالیتی مستقیم نسبت به خود سلول عمل مینمایند و چندین مسیر بازسازی برای ماکرو مولکولهای بزرگ، بای پس ضایعه و مقاومت یا آپوپتوزیس فراهم میسازد. مشخصههای اصلی مربوط به پاسخ کلی القا ژنهای متعدد، آسیب به چرخه سلولی و بازدارندگی از تقسیم سلولی است.
نقاط وارسی آسیب دیانای
بعد از آسیب وارد شدن به دیانای، نقاط وارسی چرخه سلولی فعال میگردد. فعال شدن نقاط کنترل منجر به توقف چرخه سلولی میشود و به سلول زمانی را برای بازسازی آسیب قبل از ادامه دادن فعالیت تقسیم میدهد. نقاط وارسی آسیب واردشده به دیانای در مرزهای گام نخستین رشد/مرحله اس و گام دومین رشد/مرحلهام رخ میدهد. در داخل نقطه کنترل اس نیز وجود دارد. فعال شدن نقطه کنترل با کمک دو کیناز ATM و ATR صورت میگیرد. ATM به شکستگیهای رشته دوگانه دیانای و توزیع در ساختار کروموزوم پاسخ میدهد، در حالی که ATR به صورت ابتدایی به توقف دوشاخه همتاسازی پاسخ میدهد. این کینازها مناطق هدف را در نظر دارند و در نهایت منجر به توقف چرخه سلولی میشوند. کلاسی از پروتئینهای واسطه شامل BRCA1, MDC1 و 53BP1 نیز تشخیص داده میشوند. این پروتئینها به نظر میرسد که برای انتقال سیگنال فعالکننده نقطه کنترل به پروتئینهای بخشهای پایینی مورد نیاز است.
مهمترین اهداف ATM و ATR در حرکت رو به پایین پی۵۳ است چنانچه برای القای خزان یاختهای به دنبال آسیب به دیانای مورد نیاز است. بازدارنده پی۲۱ کیناز وابسته به سایکلین به وسیله مکانیسمهای وابسته p53 و مستقل p53 القا میگردد و میتواند منجر به توقف چرخه سلولی در نقاط کنترل G1/S و G2/M به وسیله غیرفعال کردن ترکیبات سایکلین/ کیناز وابسته به سایکلین شود.
پاسخ ساس (SOS) پروکاریوت
پاسخ ساس تغییر در بیان ژن در اشریشیا کلی و دیگر باکتریها در پاسخ به آسیب دیانای است. سیستم SOS پروکاریوت به وسیله دو پروتئین اصلی تنظیم میگردد: LexA و RecA. LexA همودیمر یک رپرسور رونویسی است که به توالی اپراتور پیوند یافتهاست و معمولاً اشاره به جعبههای SOS میکند. در اشرشیا کولای LexA به عنوان تنظیمکننده رونویسی تقریباً ۴۸ ژن شناخته شدهاست که شامل ژنهای lexA و recA است (۳۷). پاسخ دهنده SOS در حیطه باکتریایی در سرتاسر دنیا شناخته شدهاست اما به مقدار زیادی در برخی از راستههای باکتریایی مانند اسپیروکتها وجود ندارد. سیگنالهای سلولی بسیار رایج کار فعال نمودن پاسخ SOS را انجام میدهند که به عنوان نواحی دیانای تک رشته هستند (ssدیانای) که حاصل از توقف دو شاخههای همتاسازی یا شکستگیهای رشته دوگانه است که به وسیله دیانای هلیکاز برای جداسازی دو رشته دیانای انجام میشود. در مرحله اولیه، پروتئین RecA به ssدیانای در ATP هیدرولیز شده پیسوند مییابد که حاصل واکنش ایجاد شده به صورت فیلامنتهای RecA–ssدیانای است. فیلامنتهای RecA–ssدیانای فعالکننده فعالیت اتوپروتئازهای LexA هستند که به مقدار زیادی منجر به کلیواژ شدن دیمر LexA و تنزل توالی LexA میشوند. از دست دادن رپرسور LexA ایجادکننده و تلقیحکننده رونویسی از ژنهای SOS است و برای تلقیح سیگنالی بیشتر، بازدارندگی از تقسیم سلولی و افزایش سطوح پروتئینها مسئول برای فرایند آسیبرسانی مجاز است.
در اشرشیا کولای، جعبههای SOS دارای ۲۰ نوکلئوتید در راستای توالی نزدیک به شناسانگر با ساختار واروخوانهای و درجه بالای تبدیل توالی هستند. در بقیه کلاسها و راستهها، توالی مربوط به جعبههای SOS به صورت قابل توجهی با طول متفاوت و ترکیب گوناگون تغییر مییابد اما این معمولاً به مقدار زیادی مورد حفاظت قرار میگیرد و یکی از قویترین سیگنالهای کوتاه در ژنوم است (۳۸). مقدار زیاد اطلاعات جعبههای SOS اجازه پیوندهای مختلف LexA را نسبت یا شناسانگرهای مختلف را میدهد و برای زمانبندی پاسخ SOS نیز مجاز هستند. ژنهای بازسازیکننده ضایعه در ابتدای پاسخ SOS تلقیح میشوند. به عنوان مثال خطا در پلیمرازهای ترنسلیژن UmuCD'2 (همچنین دیانای پلیمراز V نام دارد) بعداً تنظیم و دستهبندی نهایی را ایجاد میکند. در ابتدا دیانای آسیب دیده بازسازی میگردد یا با استفاده از پلیمراز یا از طریق ترکیب شدن مجدد بای پس میشود، مقدار دیانای تکرشتهای در سلولها کاهش مییابد، در نتیجه مقدار فیلامنتهای RecA فعالیت کلیواژ همودیمر LexA را کاهش میدهد که سپس به جعبههای SOS در نزدیکی شناساگرها پیوند مییابد.
پاسخ رونویسی یوکاریوتی به آسیب دیانای
سلولهای یوکاریوتی در معرض عوامل آسیبرسان به دیانای نیز فعالکننده مسیرهای دفاعی مهم توسط پروتئینهای مختلفی هستند که در بازسازی دیانای ، کنترل نقاط کنترل چرخه سلولی، تراکم و تنزل پروتئین نقش دارند. چنین ژنومی گسترش دهنده پاسخ رونویسی است که بسیار پیچیده است و به مقدار زیادی تنظیمکننده است، بنابراین اجازه تطابق پاسخ کلی را نسبت به آسیب میدهد. در معرض قرارگیری مخمر ساکارومایسس سرویزیه نسبت به عوامل آسیبرسان به دیانای منجر به ایجاد هم پوشانی میگردد اما به صورت پروفیلهای رونویسی مجزایی این کار انجام میشود. تشابه با پاسخ شوک زیستمحیطی نشلان دهنده این است که مسیری برای پاسخ تنشی کلی در سطح فعال ساختن رونویسی وجود دارد. برعکس، انواع مختلف سلولهای انسانی به آسیب به صورت مختلفی پاسخ میدهند که خود نشان دهنده غیاب و نبود پاسخ کلی در این زمینه است. تفسیر احتمالی برای این تفاوت بین مخمر و سلولهای انسانی ممکن است در ارتباط با هتوژنز بودن سلولهای پستانداران باشد. در حیوانات انواع مختلف سلولها در بین اندامهای مختلف وجود دارد که دارای حساسیتهای مختلفی به آسیب دیانای هستند.
در پاسخ کلی به آسیب دیانای میتوان بیان شدن چندین ژن را به عنوان مسئولی برای بازسازی بعد از تکثیر، ترکیب شدن مجدد همولوگها، بازسازی نوکلئوتید، نقاط کنترل آسیب دیانای، فعال شدن رونویسی، ژنهای کنترلکننده تخریب mRNA و بیشتر موارد دیگر بیان کرد. مقدار زیادی از آسیب وارد شده به سلول آن را رها میسازند: در این حالت آپوپتیز و مرگ رخ میدهد یا این که با یک ژنوم تغییر یافته زنده میماند. افزایش در مقاومت به آسیب میتواند منجر به افزایش مقدار حیات گردد که اجازه تجمع بیشتری را به جهشها میدهد. مخمر Rev1 و پلیمراز انسانی ηاعضای خانواده پلیمراز دیانای ترنسلیژن Y هستند که در طول پاسخ کلی به آسیب دیانای وجود دارند و مسئول ارتقا جهشها در طول پاسخ کلی به آسیب دیانای در یوکاریوتها هستند.
بازسازی دیانای و پیر شدن
اثرات پاتولوژیکی بازسازی دیانای ضعیف

حیوانات مورد آزمایش با کمبودهای ژنتیکی در بازسازی دیانای اغلب طول عمر کوتاه تری دارند و احتمال ابتلا به سرطان در آنها افزایش مییابد. به عنوان مثال موش دارای نقش در زمینه مسیر NHEJ و مکانیسمهای حفاظت تلومر است که بر روی انف تأثیر دارد و منجر به ایجاد عفونت میگردد و در نتیجه طول عمر کوتاهتر از انواع موشهای وحشی دیگر خواهد بود. در روشی مشابه، موش دارای نقص در بازسازی اصلی و پروتئین رونویسی فاقد دیانای هلیکاز برای دوران پیش از بلوغ است و در نتیجه این امر طول عمر را کوتاه میکند. به هر حال، هر کمبود در بازسازی دیانای دقیقاً اثرات پیشبینی شده را ایجاد نمیکند، موش دارای نقص در مسیر NER طول عمر کوتاه تری را بدون انطباق با مقادیر بیشتر جهش نشان میدهد. اگر میزان دیانای آسیب دیده بیش از ظرفیت سلول برای بازسازی آن باشد در این صورت تجمع خطاها میتواند منجر به آپوپتیز یا سرطان شود. بیماریهای وراثتی در ارتباط با عملکرد بازسازی دیانای آسیب دیده در پیش از سن بلوغ رخ میدهد و منجر به افزایش سرطان میشود (بخش زیر را ببینید). از سوی دیگر، ارگانیسمهایی با سیستمهای بازسازی دیانای پیشرفته مانند Deinococcus radiodurans به مقدار بیشتری مقاوم به تشعشع هستند و مقاومت قابل توجهی نسبت به شکستگی رشته دوگانه تحت تأثیر واپاشی هستهای نشان میدهند که احتمالاً ناشی از افزایش کارایی بازسازی دیانای به خصوص NHEJ است.
افزایش طول عمر و محدود کردن کالری

تعدادی از ژنهای فردی به عنوان تغییر دهندههای مدت زمان زندگی در بین ارگانیسمها شناخته شدهاند. اثرات مربوط به این ژنها به مقدار زیادی بستگی به محیط دارد و به صورت خاصی بستگی به رژیم غذایی ارگانیسم دارد. محدود کردن کالری منجر به افزایش طول عمر در انواع گوناگونی از ارگانیسمها میشود که احتمالاً از طریق تأثیر مواد غذایی و کاهش میزان دگرگشت است. مکانیسمهای مولکولی که محدودکننده هستند منجر به افزایش طول عمر میشوند که تاکنون مشخص نشدهاند (منبع ۴۴را برای بحث بیشتر ببینید)، به هر حال رفتار بیشتر ژنها شناخته شدهاست که در ارتباط با بازسازی دیانای و تغییر در شرایط محدود بودن کالری است.
به عنوان مثال افزایش در مقدار چندی ژن مربوط به ژن SIR-2 منجر به تنظیم بستهبندی دیانای در کرمهای لولهای Caenorhabditis elegans میشود که میتواند بهطور معناداری طول عمر را افزایش دهد. همولوگ SIR-2 به عنوان تلقیحکننده فاکتورهای بازسازی دیانای شناخته شدهاست که شامل NHEJ است، فعالیتی که منجر به ارتقای شرایط محدود ساختن کالری میشود. محدود ساختن کالری به مقدار زیادی در ارتباط با مقدار بازسازی باز در دیانای هسته جوندگان است اگر چه اثرات مشابه در دیانای میتوکندری مشاهده نشدهاست.
این جالب توجه است که اشاره کنیم که ژن AGE1- در C. elegans به عنوان ایفاکتوری از مسیرهای بازسازی دیانای است که به مقدار زیادی بر روی افزایش طول عمر تحت شرایط تغذیه آزاد اثر دارد اما منجر به کاهش تکثیر تحت شرایط محدود شدن کالری میشود. این مشاهده از تئوری چندنمودی (پلیوتروپی) در ارتباط با پیر شدن بیولوژیکی حمایت میکند که از این رو پیشنهاد میگردد که ژنهای مؤثر در اوایل طول عمر میتواند در پایان عمر به صورت ناموثر عمل نمایند.
پزشکی و بازسازی مدولاسیون دیانای
ارثی بودن بیماریهای بازسازی دیانای
مشکلات در ارتباط با مکانیسم NER مسئول بیماریهای مختلف ژنتیکی هستند که شامل موارد زیر هستند:
- گزرودرما پیگمنتوزوم: حساسیت به نور خورشید / UV، منجر به افزایش بروز سرطان پوست و پیری زودرس میشود.
- سندرم کوکایین: حساسیت به UV و مواد شیمیایی
- Trichothiodystrophy: پوستهای حساس، مو و ناخن شکننده
اختلالات ذهنی اغلب در ارتباط با دو بیماری اخیر است از این رو افزایش در آسیبپذیری سلولهای عصبی پیشرفته پیشنهاد میگردد.
- سندروم ورنر: پیر شدن قبل از رسیدن به سن بلوغ و متوقف شدن رشد.
- سندروم بلوم: قرار گرفتن زیاد در معرض نور خورشید، افزایش شیوع سرطان (به خصوص لوسمی)
- تلانژکتازی آتاکسی: حساسیت به پرتوهای یونیزان و برخی مواد شیمیایی عوامل
همه بیماریهای بالا اغلب پیری زودرس سگمنتال (شتاب در بیماری پیری) نام دارد زیرا قربانیهای این بیماری زود پیر میشوند و از بیمارهای مرتبط با پیری در سن جوانی به صورت غیرطبیعی رنج میبرند در حالی که همه نشانههای پیری را ندارند.
بیماریهای دیگر در ارتباط با کاهش عملکرد بازسازی دیانای، شامل آنمی فانکونی، سرطان پستان ارثی و سرطان روده بزرگ ارثی است.
بازسازی دیانای و سرطان
به دلیل محدودیتهای وراثتی در زمینه بازسازی دیانای، اگر انسانها به اندازه کافی عمر نمایند در این صورت آنها ممکن است گاهی اوقات مبتلا به سرطان شوند. حداقل ۳۴ جهش در ژن بازسازی دیانای انسان به صورت ارثی وجود دارد که خطر ابتلا به سرطان را افزایش میدهد. بیشتر این جهشها منجر به بازسازی دیانای به صورت کمتر مؤثر در مقایسه با حالت نرمال میشوند. به صورت خاصی، سرطان کولورکتال غیر پولیپوزیس ارثی یا HNPCC به مقدار زیادی در ارتباط با جهشهای خاصی در روش بازسازی عدم انطباق دیانای است. BRCA1 و BRCA2 دو ژن معروف هستند که جهشهای صورت گرفته در آنها به مقدار زیادی امکان ابتلا به به سرطان را افزایش میدهد که در ارتباط با تعداد زیادی از مسیرهای بازسازی دیانای به خصوص NHEJ و ترکیب شدن همولوگها است.
فرایندهای درمان سرطان مانند شیمیدرمانی و پرتودرمانی از تمام ظرفیت سلول برای بازسازی دیانای آسیب دیده که منجر به مرگ سلول میشود استفاده میکنند. سلولهایی که به سرعت تقسیم میگردند معمولاً ایجادکننده سرطان هستند و به مقدار زیادی تحت تأثیر قرار دارند. اثرات جانبی این است که دیگر سلولهای غیر سرطانی نیز به سرعت تقسیم میگردند مانند سلولهای مولد در روده، پوست و سیستم خون ساز تحت تأثیر قرار میگیرند.
در درمانهای جدید برای سرطان تلاش شدهاست تا مکان آسیب دیانای در سلولها و بافتها تنها در ارتباط با سرطان و با کمک ابزارهای پزشکی یا بیوشیمیایی تشخیص داده شود.
دشواریهای بازسازی اپیژنیک دیانای در سرطان
به صورت طبقهبندی شده سرطان به عنوان مجموعهای از بیماریهایی شناخته شدهاست که حاصل پیشرفت ژنتیکی غیرطبیعی است که شامل جهشهایی در ژنهای ایجادکننده تومور و چیدمانهای کوروموزومی است. به هر حال مشخص شدهاست که سرطان نیز از تغییرات وراژنتیک شکل میگیرد.
تغییرات اپی ژنیک اشاره به تغییرات عملکردی آشکار در زنوم دارد که در ارتباط با تغییر توالی نوکلئوتیدی نیست. مثالهای مربوط به چنین تغییرات شامل تغییرات در دیانای متیلاسیون و تغییر هیستون، تغییرات در معماری کوروموزومی و تغییرات ایجاد شده توسط میکرو RNAها است. هر یک از این تغییرات اپی ژنیک برای تنظیم بیانَ دن ژن بدون تغییر توالی دیانای مورد استفاده قرار میگیرند. این تغییرات معمولاً از طریق تقسیم سلولی در چندین نسل سلولی باقی میمانند و میتوان از آن به عنوان جهشهای اپی (معادل با جهش) یاد کرد.
در حالی که تعداد زیادی از تغییرات اپی ژنیک در سرطانها یافت شدهاست اما تغییرات اپی ژنیک در ژنهای بازسازی دیانای منجر به کاهش بیان شدن در پروتئینهای بازسازی دیانای میشود که دارای اهمیت زیادی است. چنین تغییراتی در اوایل پیشرفت سرطان رخ میدهد و به احتمال زیاد منجر به ایجاد مشخصات ناپایدار ژنتیکی میشود.
کاهش در بیان شدن ژنهای بازسازی دیانای منجر به کمبود و نقص در بازسازی دیانای میشود. زمانی که بازسازی دیانای با اشکال مواجه میگردد در این صورت دیانای آسیب دیده در سلولها در بالاترین سطح ممکن باقی میماند و این منجر به افزایش آسیبهای حاصل از فراوانی جهشها یا اپی جهشها میشود. مقادیر جهش اساساً منجر به افزایش سلولهای دفاعی در بازسازی غیر انطباقی دیانای یا بازسازی ترکیب شدن مجدد همولوگها یا HRR میشود. چیدمانهای مجدد کروموزومی نیز منجر به افزایش HRR سلولهای دفاعی میشود.
سطوح بالاتر دیانای آسیب دیده نه تنها منجر به افزایش جهشها نمیشود بلکه منجر به افزایش اپی جهش میشود. در طول بازسازی شکستگیهای رشته دوگانه دیانای یا بازسازی دیگر دیانایهای آسیب دیده، به صورت نیمه کاملی مکانهای بازسازی مشخص است که میتواند منجر به توقف ژنهای اپی ژنیک نیز شود.
نقص در بیان شدن بازسازی پروتئینهای دیانای به دلیل جهش ارثی میتواند منجر به افزایش خطر سرطان شود. افراد با مشکلات وراثتی در هر ۳۴ ژن بازسازی یافته (مقاله در مورد بیماریهای مربوط به کمبود بازسازی دیانای را ببینید) احتمال افزایش ابتلا به سرطان را دارند. با برخی از مشکلاتی ذکر شده احتمال ابتلا به سرطان در طول عمر ۱۰۰ درصد خواهد شد (برای مثال جهشهای پی53). به هر حال برخی از جهشهای ژرملین تنها منجر به ابتلا به سرطان به میزان ۱ درصد میشوند.
فراوانی اپی جهشها در ژن ای بازسازیکننده دیانای

مشکلات و نقصهای مربوط به آنزیمهای بازسازی دیانای به صورت اتفاقی حاصل از جهشهایی در ژن بازسازی دیانای است اما به مقدار زیادی توسط تغییرات اپی ژنیک شکل میگیرد که منجر به کاهش یا توقف بیان شدن ژنهای بازسازی دیانای میشوند. به عنوان مثال، زمانی که ۱۱۳ توالی مربوط به افراد دارای سرطان روده بزرگ مورد آزمون قرار گرفت تنها ۴ نفر فاقد جهش در ژن بازسازی دیانای با نام MGMT بودند، در حالی که اکثر آنها دارای بیان کاهش یافتهای از MGMT بودند که ناشی از متیلی شدن ناحیه شناساگر MGMT بود (یک تغییر اپی ژنیک). در پنج مطالعه مختلف نشان داد که ۴۰ تا ۹۰ درصد از افراد مبتلا به سرطان روده بزرگ دارای بیان کاهش یافتهای از MGMT بودند که این امر ناشی از متیلی شدن ناحیه شناساگر MGMT بود.
به صورت مشابهی به غیر از ۱۱۹ مورد سرطان روده بزرگ دارای نقص بازسازی غیر انطباقی که فاقد ژن بازسازی دیانای با نام بیان شدن PMS2 بودند، PMS2 در ۶ مورد نقص داشت که ناشی از جهشها در ژن PMS2 بود، در حالی که در ۱۰۳ مورد بیان شدن ژن PMS2 به دلیل جفت شدن MLH1 به دلیل شناساگر متیل شدن نقص داشت (پروتئین PMS2 در غیاب MLH1 ناپایدار است). در ۱۰ مورد دیگر، از دست دادن بیان PMS2 به احتمال زیاد ناشی از بیان شدن بیش از حد اپی ژنتیک microRNA, miR-155 بود که به مقدار کمتری به تنظیم MLH1 میپرداخت.
در مثالهای دیگر (بخش مربوط به اپی ژنتیکهای بازسازی دیانای را در سرطان ببینید) مشکلات اپی ژنتیک به صورت مکرر به میزان ۱۳ تا ۱۰۰ درصد برای ژنهای بازسازی دیانای به شرح BRCA1, WRN, FANCB, FANCF, MGMT, MLH1, MSH2, MSH4, ERCC1, XPF, NEIL1 و ATM مشاهده شدند. این مشکلات و نقص اپی ژنتیکی در سرطانهای مختلفی رخ میدهد (برای مثال سینه، تخمدان، روده بزرگ، سر و گردن). دو یا سه نقص در بیان ERCC1, XPF یا PMS2 به صورت همزمان در اکثر ۴۹ مورد مبتلا به سرطان کولون رخ داده است که به وسیله فاسیتا و همکاران مورد ارزیابی قرار گرفتهاست.
نمودار موجود در این بخش برخی از عوامل آسیب به دیانای را نشان میدهد، مثالهای مربوط به ضایعات دیانای و مسیرهای مربوط به این آسیبهای وارده به دیانای را ببینید. حداقل ۱۶۹ آنزیم وجود دارد که به صورت مستقیم برای بازسازی دیانای یا اثر بر روی فرایند بازسازی دیانای به کار برده میشوند. از بین اینها ۸۳ مورد به صورت مستقیم در ۵ نوع فرایند بازسازی دیانای به کار رفتهاند که در نمودار نشان داده شدهاست. ژنهای مطالعه شده دیگر نیز در راس فرایندهای بازسازی در نمودار نشان داده شدهاند. چنانچه توسط ژنهای بازسازی ژن به صورت قرمز رنگ نشان داده شدهاست، بیشتر این ژنها در این مسیرهای بازسازی توسط مکانیسمهای اپی ژنتیکی تنظیم میگردند و اینها به صورت مکرری کاهش مییابند یا این که در سرطانهای مختلف متوقف میگردند. دو مقاله بررسی شده، و دو مقاله گستردهتر در این زمینه بیشتر این کمبودها را در زمینه بازسازی اپی ژنتیک را نشان میدهد.
این نشان دهنده تغییرات اپی ژنیک در ژنهای بازسازی دیانای است که نقش اصلی در ابتلا به سرطان را ایفا میکنند.
بازسازی دیانای و سیر تکاملی آن
فرایندهای اصلی بازسازی دیانای به مقدار زیادی هم در پروکاریوتها و هم در یوکاریوتها و حتی در میان فاژها حفاظت شدهاست (ویروسهایی که عفونت باکتریایی دارند) به هر حال بیشتر ارگانیسمهای پیچیده با ژنومهای بسیار پیچیده منطبق با مکانیسمهای بازسازی بسیار پیچیده هستند. توانایی مربوط به تعداد زیادی از موتیفهای ساختاری پروتئین برای کاتالیز واکنشهای شیمیایی نقش معناداری را در ارتقا مکانیسمهای بازسازی در طول سیر تکاملی ایفا میکند. برای بررسی فرضیه با جزئیات بیشتر در ارتباط با سیر تکاملی فرایند بازسازی دیانای منبع را ببینید.
سنگوارهها نشان میدهند که آغاز زندگی بر روی نقاط سیاره در برخی مکانها در طول دوران پرکامبرین به صورت تک سلولی بودهاست، اگر چه پیدایش اولیه حیات نیز اکنون دقیقاً مشخص نیست. اسید نوکلئیک کل اطلاعات ژنتیکی را در سرتاسر دنیا کدگذاری میکنند و مکانیسمهای بازسازی دیانای مورد نیاز را تعیین میکنند که شکل اصلی آنها توسط همه شکلهای گسترش یافته زندگی از اجداد گذشته ما به ارث رسیدهاست. با افزایش یافتن میزان اکسیژن در اتمسفر به دلیل فوتوسنتز ارگانیسمها، و به علاوه حضور رادیکالهای آزاد آسیبرسان در سلول به دلیل فوسفوریلاسیون فعال، بررسی سیر تکاملی مکانیسمهای بازسازی دیانای ضروری است که به صورت خاصی برای مقابله با استرس ناشی از اکسیداتیو ضروری است.
میزان تغییر سیر تکاملی
براساس برخی از رخدادها، دیانای آسیب دیده بازسازی نمییابد با این که توسط مکانیسم خطایی بازسازی مییابد که منجر به ایجاد تغیر توالیهای اصلی میشود. زمانی که این امر رخ میدهد، جهشها ممکن است نسبت به ژنومهای سلولهای فرزندی رخ دهد. باید چنین رخدادهایی در سلول در حالت تکثیر رخ دهد کا گاهی اوقات تولید گامت میکند، جهش دارای توان بالقوه برای انتقال قدرت باروری است. میزان فرگشت در یک گونه خاص (یا در یک ژن خاص) به صورت تابعی از میزان جهش است. در نتیجه، میزان و دقت مکانیسمهای بازسازی دیانای بر روی فرایند سیر تکاملی تغییر تأثیر دارد.
- ترجمه از ویکیپدیا انگلیسی