
| Negarnaviricota | |
|---|---|
| |
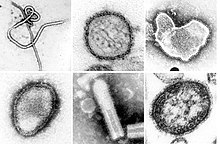
| A montage of transmission electron micrographs of some viruses in the phylum Negarnaviricota. Not to scale. Species from left to right, top to bottom: Zaire ebolavirus, Sin Nombre orthohantavirus, Human orthopneumovirus, Hendra henipavirus, an unidentified rhabdovirus, Measles morbillivirus. | |
|
طبقهبندی ویروسها | |
| (طبقهبندینشده): | ویروس |
| Realm: | ریبوویرا |
| شاخه: | Negarnaviricota |
| Subtaxa | |
| مترادف | |
|
Negative-sense RNA viruses (NSVs) | |
آرانای ویروسهای سوی منفی (Negative-sense RNA viruses) گروهی از ویروسهای مرتبط هستند که دارای ژنومهای تکرشتهای سوی منفی و حاوی اسید ریبونوکلئیک هستند. آنها ژنومهایی دارند که بهعنوان رشتههای مکمل عمل میکنند که از آن، آرانای پیامرسان توسط آنزیم ویروسی آرانای پلیمراز وابسته به آرانای (RDRP) سنتز میشود. این آنزیم در طول تکثیر ژنوم ویروسی، یک آنتی ژنوم با سوی مثبت را سنتز میکند که از آن بهعنوان الگویی برای ایجاد آرانای با ژنوم سوی منفی استفاده میکند. آرانای ویروسهای سوی منفی تعدادی ویژگی دیگر را نیز به اشتراک میگذارند: اکثر آنها حاوی یک پوشش ویروسی هستند که کپسید را احاطه کردهاست، که ژنوم ویروس را در بر میگیرد، ژنومهای این ویروسها از نوع آرانای تکرشتهای معمولاً خطی هستند.
آرانای ویروسهای سوی منفی، شاخه Negarnaviricota را در قلمرو Orthornavirae و قلمرو ریبوویرا تشکیل میدهند. آنها از یک نیای مشترک که یک آرانای ویروس دو رشتهای بود، به وجود میآیند، و آنها را خواهری از رئوویروسها میدانند که آرانای ویروسهای دورشتهای هستند. در داخل این شاخه، دو شاخهٔ اصلی وجود دارد که دو زیرشاخه را تشکیل میدهند: هاپلوویریکوتینا، که اعضای آن عمدتاً غیرقطعی هستند و یک آردیآرپی را رمزگذاری میکنند که کلاهکهای روی آرانای پیامرسان را سنتز میکند و Polyploviricotina، که اعضای آن قطعهبندی شدهاند و یک آردیآرپی را کد میکند که کلاهکها را از آن جدا میکند. آرانایهای پیامرسان میزبان در مجموع شش کلاس در این شاخه به رسمیت شناخته شدهاست.
آرانای ویروسهای سوی منفی ارتباط نزدیکی با بندپایان دارند و میتوانند بهطور غیررسمی بین آنهایی که برای انتقال به بندپایان متکی هستند و ویروسهایی که از ویروسهای بندپایان منشأ میگیرند اما اکنون میتوانند بدون کمک بندپایان در مهرهداران تکثیر شوند، تقسیم شوند. آرانای ویروسهای سوی منفی برجسته بندپایان شامل ویروس تب دره ریفت و ویروس پژمردگی لکهدار گوجهفرنگی هستند. آرانای ویروسهای سوی منفی قابل توجه مهرهداران شامل ویروس ابولا، هانتاویروسها، ویروسهای آنفولانزا، ویروس تب لاسا و ویروس هاری هستند.
ویژگیها
ژنوم

همه ویروسهای Negarnaviricota، ویروسهایی با سوی منفی و تکرشتهای آرانای (اساس آرانای) هستند. آنها دارای ژنومهای ساخته شده از آرانای هستند که به جای دو رشتهای، تک هستند. ژنوم آنها دارای سوی منفی است، به این معنی که آرانای پیامرسان میتواند بهطور مستقیم از ژنوم توسط آنزیم ویروسی آر ان ای پلیمراز وابسته به آرانای (آر دی آر پی)، که آر ان ای replicase نیز نامیده میشود، که توسط همه ویروسهای -اس اس آر ان ای کدگذاری میشود، سنتز شود. به استثنای ویروسهای جنس Tenuivirus و برخی از خانواده Chuviridae، همه ویروسهای −اس اس آر ان ای ژنومهای خطی دارند تا دایرهای، و ژنومها ممکن است قطعهبندی یا غیرقطعی باشند. همه ژنومهای اس اس آر ان ای حاوی تکرارهای معکوس انتهایی هستند که توالیهای نوکلئوتیدی پالیندرومیک در هر انتهای ژنوم هستند.
همانندسازی و رونویسی
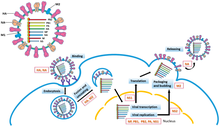
همانندسازی ژنوم آرانای ویروسهای دورشتهای توسط آر دی آر پی اجرا میشود، که همانندسازی را با اتصال به یک توالی رهبر در انتهای ۳' (معمولاً «سه انتهای اصلی» تلفظ میشود) ژنوم آغاز میکند. سپس آر دی آر پی از ژنوم سوی منفی به عنوان الگویی برای سنتز آنتی ژنوم با حس مثبت استفاده میکند. هنگام تکثیر آنتی ژنوم، آر دی آر پی ابتدا به دنباله تریلر در انتهای ۳' آنتی ژنوم متصل میشود. پس از آن، آر دی آر پی تمام سیگنالهای رونویسی روی آنتی ژنوم را نادیده میگیرد و یک کپی از ژنوم را در حالی که از آنتی ژنوم به عنوان الگو استفاده میکند سنتز میکند. همانندسازی زمانی انجام میشود که ژنوم داخل نوکلئوکپسید است و آر دی آر پی کپسید را آشکار میکند و در طول همانندسازی در طول ژنوم جابهجا میشود. همانطور که توالیهای نوکلئوتیدی جدید توسط آر دی آر پی سنتز میشوند، پروتئینهای کپسید مونتاژ میشوند و آر ان ای ویروسی تازه تکثیر شده را محصور میکنند.
رونویسی آرانای پیامرسان از ژنوم از همان الگوی جهتی پیروی میکند که تولید آنتی ژنوم است. در دنباله رهبر، آر دی آر پی یک آر ان ای پیشرو تری فسفات با ۵ 'انتها (معمولاً «پنج انتهای اول» تلفظ میشود) را سنتز میکند و در مورد زیر شاخه Haploviricotina ، انتهای ۵ را میپوشاند یا در مورد زیر شاخه. Polyploviricotina، کلاهکی را از آرانای پیامرسان میزبان میرباید و آن را به آرانای پیامرسان ویروسی متصل میکند تا آرانای پیامرسان توسط ریبوزومهای سلول میزبان ترجمه شود.
پس از پوشاندن آرانای پیامرسان، آر دی آر پی رونویسی را در سیگنال شروع ژن آغاز میکند و بعداً با رسیدن به سیگنال پایان ژن رونویسی را پایان میدهد. در پایان رونویسی، آر دی آر پی یک دم پلی آدنیله متشکل از صدها آدنین در انتهای ۳' آرانای پیامرسان را سنتز میکند، که ممکن است روی دنبالهای از اوراسیلها انجام شود. پس از ساخت دم پلی (A)، آرانای پیامرسان توسط آر دی آر پی آزاد میشود. در ژنومهایی که بیش از یک بخش قابل رونویسی را رمزگذاری میکنند، آر دی آر پی میتواند به اسکن دنبالهٔ شروع بعدی ادامه دهد تا به رونویسی ادامه دهد.
برخی از آرانای ویروسهای سوی منفی امبیسنس هستند، به این معنی که هم رشتهٔ ژنومی منفی و هم آنتیژنوم مثبت بهطور جداگانه پروتئینهای مختلف را کد میکنند. به منظور رونویسی ویروسهای امبیسنس، دو دور رونویسی انجام میشود: اول، ام آر ان ای بهطور مستقیم از ژنوم تولید میشود. دوم، آرانای پیامرسان از آنتی ژنوم ایجاد میشود. همه ویروسهای ambisense دارای ساختار حلقهای هستند که پس از رونویسی ام آر ان ای پروتئین، رونویسی را متوقف میکنند.
مورفولوژی

آرانای ویروسهای سوی منفی حاوی یک کمپلکس ریبونوکلئوپروتئینی متشکل از ژنوم و یک آر دی آر پی متصل به هر بخش از ژنوم است که توسط یک کپسید احاطه شدهاست. کپسید از پروتئینهایی تشکیل شدهاست که ساختار چین خورده آنها شامل پنج مارپیچ آلفا در لوب پایانه-N (موتیف 5-H) و سه مارپیچ آلفا در لوب پایانه-C (موتیف 3-H) است. در داخل کپسید، ژنوم بین این دو نقوش قرار گرفتهاست. به استثنای خانواده Aspiviridae، آرانای ویروسهای سوی منفی حاوی یک پوشش ویروسی خارجی هستند، نوعی غشای لیپیدی که کپسید را احاطه کردهاست. شکل ذره ویروس که ویریون نامیده میشود، آرانای ویروسهای سوی منفی متفاوت است و ممکن است رشتهای، پلئومورفیک، کروی یا لولهای باشد.
فرگشت
تقسیمبندی ژنوم یک ویژگی برجسته در میان بسیاری از ویروسهای -اس اس آر ان ای است، و ویروسهای -اس اس آر ان ای از داشتن ژنوم با یک بخش، عادی برای اعضای راسته مونونگاویرالس، تا ژنومهایی با ده بخش، مانند ویروس تیلاپیا تیلاپین، متغیر است. هیچ روند مشخصی در طول زمان وجود ندارد که تعداد بخشها را تعیین کند، و به نظر میرسد تقسیمبندی ژنوم در میان آرانای ویروسهای سوی منفی یک ویژگی انعطافپذیر باشد زیرا بهطور مستقل در موارد متعدد فرگشت یافتهاست. اکثر اعضای زیر شاخه هاپلوویریکوتینا غیرقطعی هستند، در حالی که تقسیمبندی در Polyploviricotina جهانی است.
فیلوژنتیک

تجزیه و تحلیل فیلوژنتیکی بر اساس آر دی آر پی نشان میدهد که آرانای ویروسهای سوی منفی از یک نیای مشترک منشأ میگیرند و احتمالاً یک کلاد خواهر از رئوویروسها هستند که ویروسهای دی اس آر ان ای هستند. در داخل شاخه، دو شاخه واضح وجود دارد که به دو زیر شاخه اختصاص داده میشوند، بر این اساس که آیا آر دی آر پی یک کلاهک را روی ام آر ان ای ویروسی سنتز میکند یا یک کلاهک را از ام آر ان ای میزبان ربوده و آن کلاه را به ام آر ان ای ویروسی متصل میکند.
در این شاخه، ویروسهای -اس اس آر ان ای که بندپایان را آلوده میکنند، ظاهراً پایه و اجداد سایر ویروسهای -اس اس آر ان ای هستند. بندپایان اغلب در گروههای بزرگ با هم زندگی میکنند، که امکان انتقال آسان ویروسها را فراهم میکند. با گذشت زمان، این باعث شدهاست که آرانای ویروسهای بندپایان سطح بالایی از تنوع را به دست آورند. در حالی که بندپایان میزبان مقادیر زیادی ویروس هستند، در مورد میزان انتقال بین گونهای ویروسهای بندپایان آرانای ویروسهای سوی منفی در بین بندپایان اختلاف نظر وجود دارد.
آرانای ویروسهای سوی منفی گیاهان و مهرهداران از نظر ژنتیکی با ویروسهای آلوده به بندپایان مرتبط هستند. علاوه بر این، بیشتر آرانای ویروسهای سوی منفی خارج از بندپایان در گونههایی یافت میشوند که با بندپایان تعامل دارند؛ بنابراین بندپایان هم به عنوان میزبان کلیدی و هم به عنوان ناقل انتقال آرانای ویروسهای سوی منفی عمل میکنند. از نظر انتقال، آرانای ویروسهای سوی منفی غیر بندپایان را میتوان بین ویروسهایی که برای انتقال به بندپایان متکی هستند و ویروسهایی که میتوانند بدون کمک بندپایان در بین مهرهداران گردش کنند، تمایز قائل شد. گروه دوم احتمالاً از اولی سرچشمه گرفتهاند و با انتقال فقط مهره داران سازگار شدهاند.
طبقهبندی
Negarnaviricota متعلق به قلمرو Orthornavirae است که شامل همه آرانای ویروسها که آر دی آر پی را کد میکنند، و قلمرو Riboviria که شامل Orthornavirae و همچنین همه ویروسهایی است که رونوشت معکوس را در پادشاهی Paraآر ان ایvirae رمزگذاری میکنند. Negarnaviricota شامل دو زیر شاخه است که شامل شش کلاس ترکیبی است که پنج تای آنها یکنواخت تا گونههای پایینتر هستند:
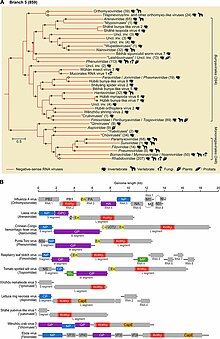
- زیر شاخه: Haploviricotina، که حاوی آرانای ویروسهای سوی منفی است که یک آر دی آر پی را رمزگذاری میکند که ساختار کلاهکی را روی آرانای پیامرسان ویروسی سنتز میکند و معمولاً ژنومهای غیرقطعی دارند.
- طبقه: Chunquiviricetes
- راسته: Muvirales
- خانواده: Qinviridae
- سرده: Yingvirus
- خانواده: Qinviridae
- راسته: Muvirales
- طبقه: Milneviricetes
- راسته: Serpentovirales
- خانواده: Aspviridae
- سرده: افیوویروس
- خانواده: Aspviridae
- راسته: Serpentovirales
- طبقه: Monjiviricetes
- کلاس: Yunchangviricetes
- راسته: Goujianvirales
- خانواده: Yueviridae
- سرده: Yuyuevirus
- خانواده: Yueviridae
- راسته: Goujianvirales
- طبقه: Chunquiviricetes
- زیر شاخه: Polyploviricotina، که حاوی آرانای ویروسهای سوی منفی است که یک آر دی آر پی را کد میکند و یک کلاهک از آرانای پیامرسان میزبان میگیرد تا بهعنوان سرپوش ام آر ان ای ویروسی استفاده کند و ژنومهای تقسیمبندی شده دارند.
- رده: Ellioviricetes
- راسته: بانیاویرالز
- رده: Insthoviricetes
- راسته: Articulavirales
- رده: Ellioviricetes
ویروسهای آر ان ای رشته منفی در سیستم طبقهبندی بالتیمور بهعنوان گروه V طبقهبندی میشوند، که ویروسها را بر اساس نحوه تولید آرانای پیامرسان آنها گروهبندی میکند و اغلب در کنار طبقهبندی استاندارد ویروس، که بر اساس تاریخچه تکاملی است، استفاده میشود؛ بنابراین گروه V و Negarnaviricota مترادف هستند.
بیماری
آرانای ویروسهای سوی منفی باعث بسیاری از بیماریهای شناخته شده میشوند. بسیاری از اینها توسط بندپایان منتقل میشوند، از جمله ویروس تب دره ریفت و ویروس پژمردگی خالدار گوجه فرنگی. در میان مهره داران، خفاشها و جوندگان ناقل رایج بسیاری از ویروسها هستند، از جمله ویروس ابولا و ویروس هاری که توسط خفاشها و سایر مهرهداران منتقل میشوند، و ویروس تب لاسا و هانتاویروسها که توسط جوندگان منتقل میشوند.ویروس آنفولانزا در بین پرندگان و پستانداران رایج است. آرانای ویروسهای سوی منفی خاص انسان شامل ویروس سرخک و ویروس اوریون هستند.
تاریخچه
بسیاری از بیماریهای ناشی از آرانای ویروسهای سوی منفی در طول تاریخ شناخته شدهاند، از جمله عفونت هانتاویروس، سرخک و هاری. در تاریخ مدرن، برخی مانند ابولا و آنفولانزا باعث شیوع بیماریهای مرگبار شدهاند. ویروس استوماتیت تاولی که برای اولین بار در سال ۱۹۲۵ جدا شد و یکی از اولین ویروسهای حیوانی مورد مطالعه قرار گرفت، زیرا میتوان آن را در کشتهای سلولی به خوبی مطالعه کرد، به عنوان یک آرانای ویروسهای سوی منفی شناسایی شد که در آن زمان منحصربهفرد بود زیرا سایر آرانای ویروسها که قبلاً شناسایی شده بودند. کشف شده سوی مثبت بودند. در اوایل قرن بیست و یکم، بیماری گاوی که توسط ویروس طاعون گاوی ایجاد میشود، پس از آبله که توسط یک دیانای ویروس ایجاد میشود، به دومین بیماری ریشهکنشده تبدیل شد.
در قرن بیست و یکم، متاژنومیکس ویروسی برای شناسایی ویروسهای موجود در محیط رایج شدهاست. برای آرانای ویروسهای سوی منفی، این امکان شناسایی تعداد زیادی از بیمهرگان، و بهویژه بندپایان، ویروسها را فراهم کرد که به ارائه بینشی از تاریخچه تکاملی آرانای ویروسهای سوی منفی کمک کرد. بر اساس تجزیه و تحلیل فیلوژنتیکی آر دی آر پی که نشان میدهد آرانای ویروسهای سوی منفی از یک جد مشترک، Negarnaviricota و دو زیر شاخه آن در سال ۲۰۱۸ تأسیس شدهاند، و در قلمرو تازه تأسیس آن زمان ریبوویریا قرار گرفتهاند.
بیشتر خواندن
- Ward, C. W. (1993). "Progress towards a higher taxonomy of viruses". Research in Virology. 144 (6): 419–53. doi:10.1016/S0923-2516(06)80059-2. PMC 7135741. PMID 8140287.